棒曲霉素(PAT)是一种剧毒真菌毒素,由于其污染水果及其衍生产品,对健康构成重大风险。 最近消除PAT的生物学策略主要集中在利用组学技术阐明拮抗微生物的分子解毒过程。 然而,目前对生物酶的快速筛选和催化机理的研究还很缺乏。 在本研究中,通过整合转录组学和分子对接,从Meyerozyma guilliermondii中快速鉴定出能够降解PAT的短链脱氢酶/还原酶(MgSDR1)。 在还原性烟酰胺腺嘌呤二核苷酸磷酸(NADPH)存在的情况下,MgSDR1在2小时内将PAT完全降解为E-ascladiol。 生物降解效率受温度、pH、酶/底物浓度、金属离子和有机试剂的影响。 值得注意的是,MgSDR1在保持梨汁关键品质参数(如颜色参数、pH、多酚氧化酶活性、维生素C含量、总酚含量、可滴定酸度和可溶性固溶物)的情况下,表现出高效的PAT降解能力。 降解过程提高了果汁的抗氧化能力,丰富了果汁中的芳香族化合物。 此外,位点定向诱变揭示了催化三联体(Ser174-Tyr188-Lys192)在MgSDR1活性中的重要作用。 本研究为PAT降解酶的筛选提供了一种有效的方法,为食品工业中真菌毒素的去除奠定了理论基础。
转录组测序分析
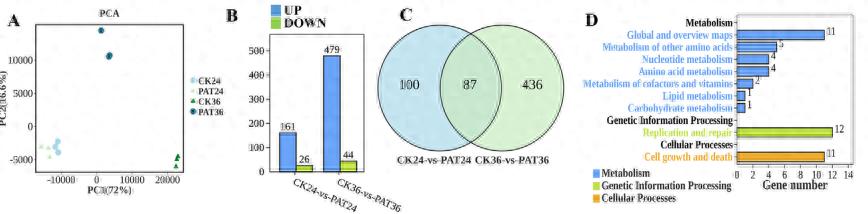
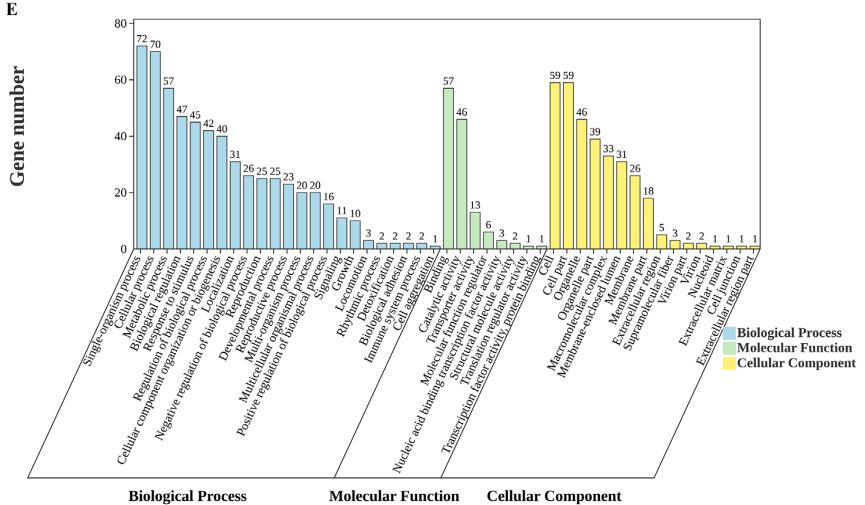
总共构建了12个文库。 图1A显示了这些文库的主成分分析(PCA),表明来自同一组的样本聚集在一起具有相似的PC1值,这解释了大部分变化(72%)。 如图1B所示,与对照组相比,处理组在24 h时有161个基因显著上调,26个基因显著下调。36 h时,处理组有479个基因显著上调,44个基因显著下调。 CK24与PAT24、CK36与PAT36之间共有87个共同基因(图1C)。KEGG注释分析这87个差异表达基因(deg)显示,它们富集于3类:代谢(7个亚类)、遗传信息处理(1个亚类)和细胞过程(1个亚类)。 氨基酸代谢、碳水化合物代谢、其他氨基酸代谢、复制和修复可能与M. guilliermondii的生长、应激反应调节和抗逆性有关(图1D)。 GO注释分析表明,这87个deg主要富集在3大类:生物过程(23个亚类)、分子功能(8个亚类)和细胞成分(16个亚类)。 细胞膜组分和细胞膜亚类可能参与了转运PAT的过程。 转运活性可能与膜的转运功能有关。 其抗胁迫活性可能与氧化活性和催化活性有关。 M. gillliermondii去除PAT可能涉及生物调控、信号传导、解毒和对刺激亚类的反应(图1E)。
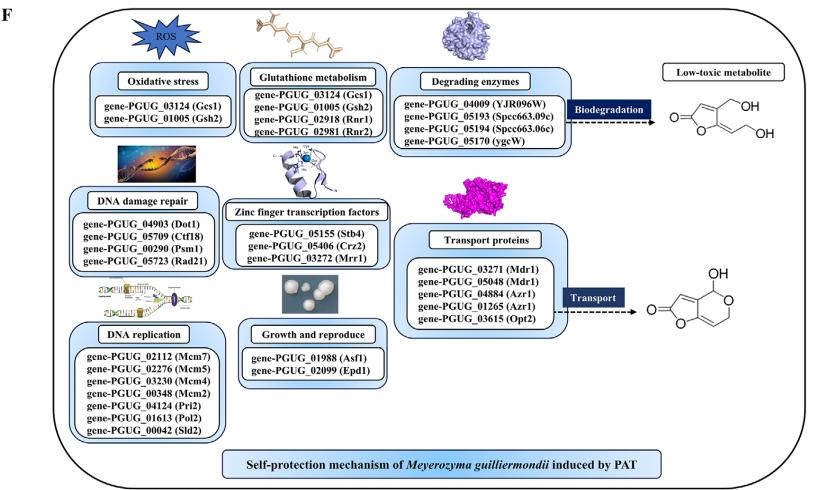
根据GO和KEGG数据库的注释信息,对deg进行进一步分类,鉴定出29个基因可能诱导PAT表达并进行自作者保护。 这些基因主要与氧化应激、谷胱甘肽(GSH)代谢、DNA损伤修复、DNA复制、生长繁殖、耐药和耐药、降解酶、锌指转录因子等相关。 这些基因的相关数据见图1F。
蛋白质与PAT的分子对接
使用ERRAT和PROCHECK软件对蛋白质结构模型的准确性进行评估 ERRAT识别不正确的非键原子接触,质量分数超过90%,证实了模型的适用性。 Ramachandran图分析表明,约90%的残基位于最有利的区域,表明在预测的三维结构中存在合适的Psi/Phi二面角。 这一发现支持了这些模型在进一步分子对接分析中的适用性。
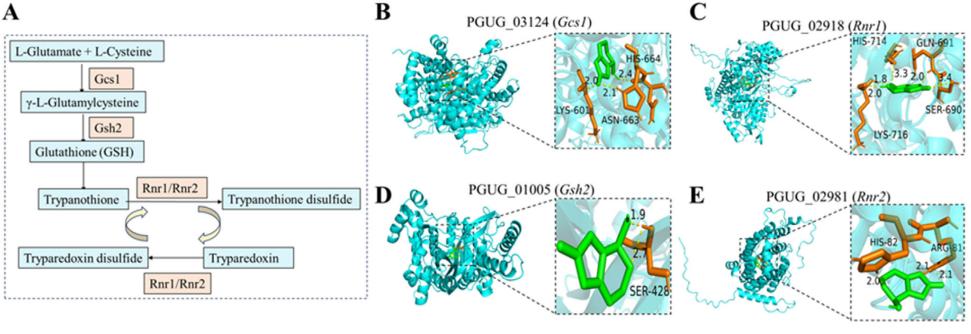
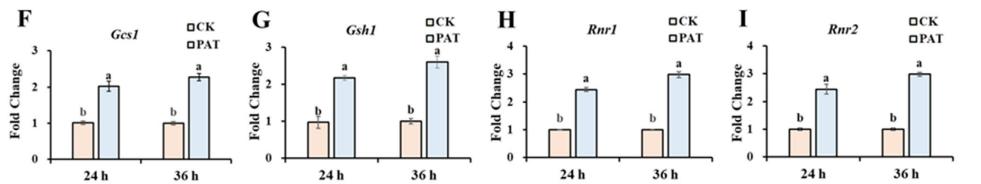
在谷胱甘肽代谢途径中,编码谷氨酸-半胱氨酸连接酶催化亚基的Gcs1、编码谷胱甘肽合成酶的Gsh2、编码核糖核苷-二磷酸还原酶亚基M1和M2的Rnr1和Rnr2四个基因显著上调(图2A)。 通过分子对接模拟了上述四种蛋白与PAT的结合。 如图2B-E所示,与GSH代谢途径相关的酶可以与PAT形成氢键,这些氢键的长度从1.8到3.4 Å不等。 此外,RT-qPCR分析显示,这四个基因的表达水平显著升高。 (图2 f i)。
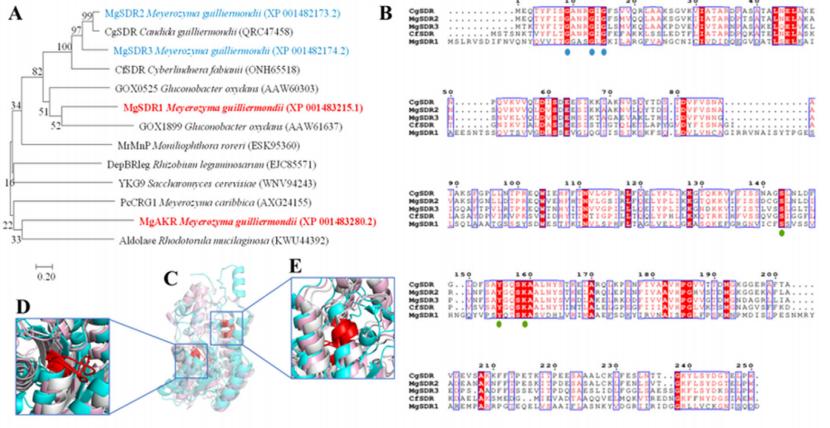
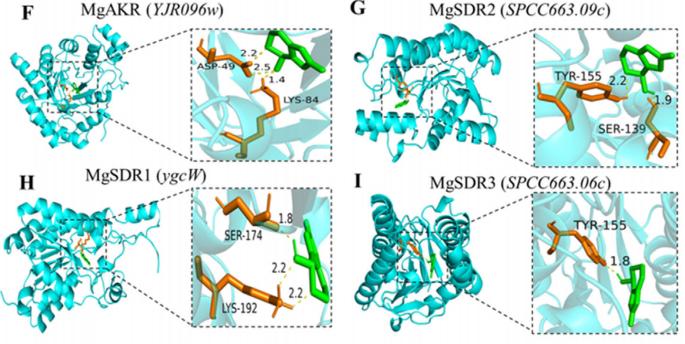
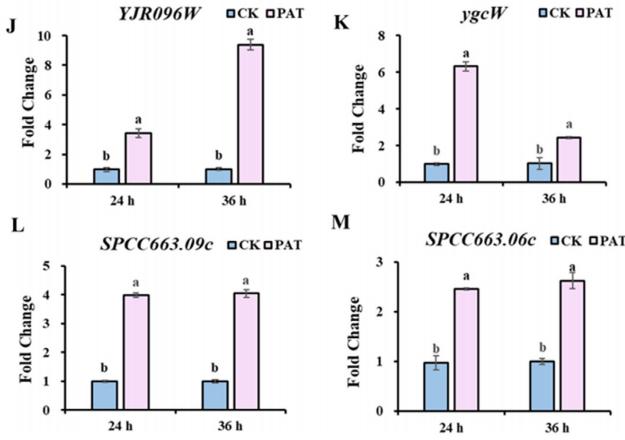
分子对接用于从DEGs中筛选具有催化和氧化还原活性的能够与PAT结合的蛋白质。 结合相关文献,共筛选出4种潜在降解酶,其编码基因均显著上调。 在潜在降解酶中,4个基因显著上调; YJR096 W编码醛酮还原酶(MgAKR), ygcW、SPCC663.09c和SPCC663.06c编码MgSDR1、MgSDR2和MgSDR3, respectively。 对这四种降解酶和文献中报道的降解酶进行系统发育树分析发现,MgAKR和MgSDR1与之前报道的降解酶具有较低的相似性,表明它们具有潜在的新颖性(图3A)。 mgakr的功能在以前的研究中已经有报道。 因此,作者的主要重点是研究MgSDR1的功能。 MgSDR1可以与SDRs家族的辅因子结合位点(GXXXGXG)和活性位点(YXXXK)对齐(图3B-E)。 如图3f - 1所示,潜在的降解酶可以通过氢键与PAT结合,这些氢键的长度从1.8到2.5 Å不等。 RT-qPCR分析显示,这四个基因的表达水平均显著升高(图3J-M)。
蛋白表达
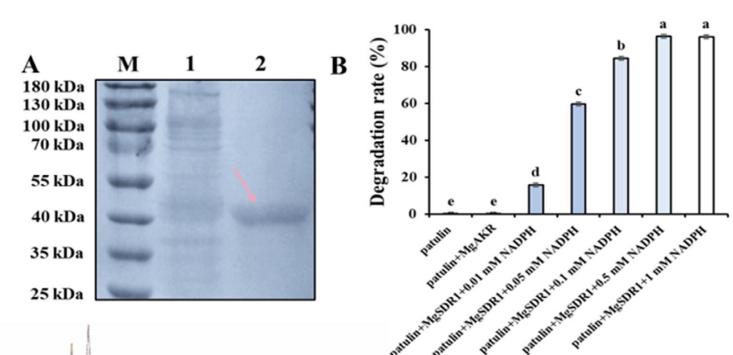
这些蛋白的表达中,带有n端6 × His-Tag的MgAKR、MgSDR1、MgSDR2和MgSDR3的分子量分别为≈38、40、36和35 kDa。原核表达结果表明,MgSDR1和MgAKR以可溶性形式表达,而MgSDR2和MgSDR3主要存在于包涵体中。在PAT上表达MgSDR1的大肠杆菌粗酶溶液优于不诱导蛋白表达的对照组的降解效果。
分子动力学模拟
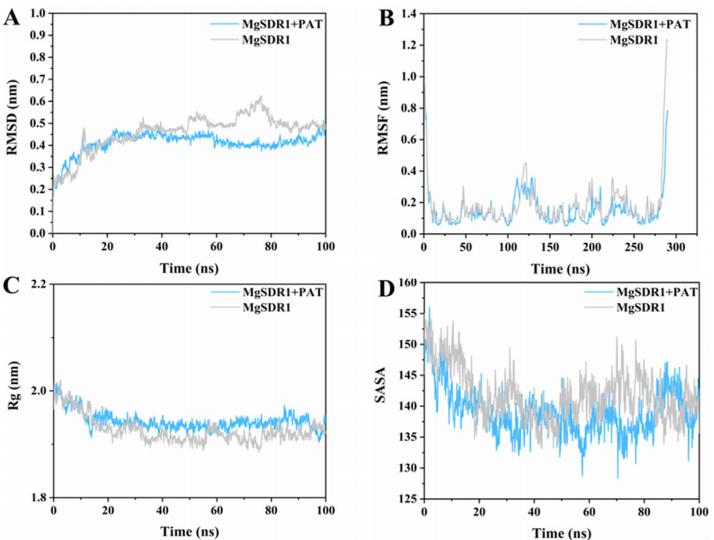
通过分子动力学模拟分析了MgSDR1-PAT配合物的构象变化和稳定性。 均方根偏差(RMSD)分析表明,MgSDR1-PAT复合物在前10ns经历了轻微的波动,主要归因于PAT结合,然后在模拟的其余部分实现稳定。 与单独的MgSDR1相比,MgSDR1PAT复合物显示出较低的RMSD,这表明MgSDR1和PAT之间存在很强的关联(图4A)。 总体而言,在100 ns模拟期间,MgSDR1- PAT复合物的均方根波动(RMSF)低于MgSDR1,这表明MgSDR1- PAT复合物表现出更大的稳定性(图4B)。 在整个模拟过程中,MgSDR1- PAT复合物和MgSDR1在溶剂可及表面积(SASA)上观察到轻微的变化。 这表明PAT结合可能在一定程度上影响MgSDR1的表面可及性,尽管总体影响很小(图4C)。 图4D显示,与MgSDR1相比,MgSDR1- PAT复合物的旋转半径(Rg)略有改变。 然而,MgSDR1-PAT复合物总体上表现出更低的Rg波动,这表明PAT结合对整体致密性的影响最小,在MgSDR1中,它可能有助于维持MgSDR1的稳定性。
NADPH浓度对MgSDR1降解PAT的影响及降解产物的验证
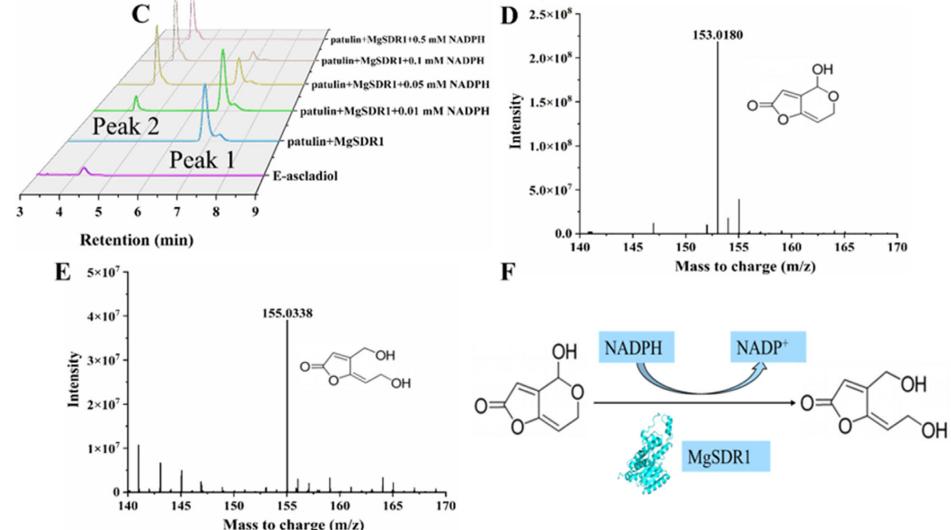
MgSDR1进行了额外的纯化以获得单个不同的目标带(图5A)。 NADPH是许多酶促反应的辅助因子。 当其浓度达到0.5 mM时,2 h时无法检测到PAT(图5B)。 此外,作者还对MgSDR1降解PAT的产物进行了分析。 高效液相色谱(HPLC)显示,PAT在6.9 min时达到峰值(峰1),在4.2 min时产生一个新的产物峰(峰2)(图5C)。 采用液相色谱-电喷雾-质谱法(LC-ESI/MS)鉴定了PAT及其降解产物的分子量。 在负离子模式下,PAT及其降解产物的分子量分别为153.0180和155.0338。 总体而言,MgSDR1将PAT转化为E-ascladiol表现出严格的还原烟酰胺腺嘌呤二核苷酸磷酸(NADPH)依赖性(图5D-F)。
反应条件、金属离子和有机试剂对MgSDR1降解PAT的影响
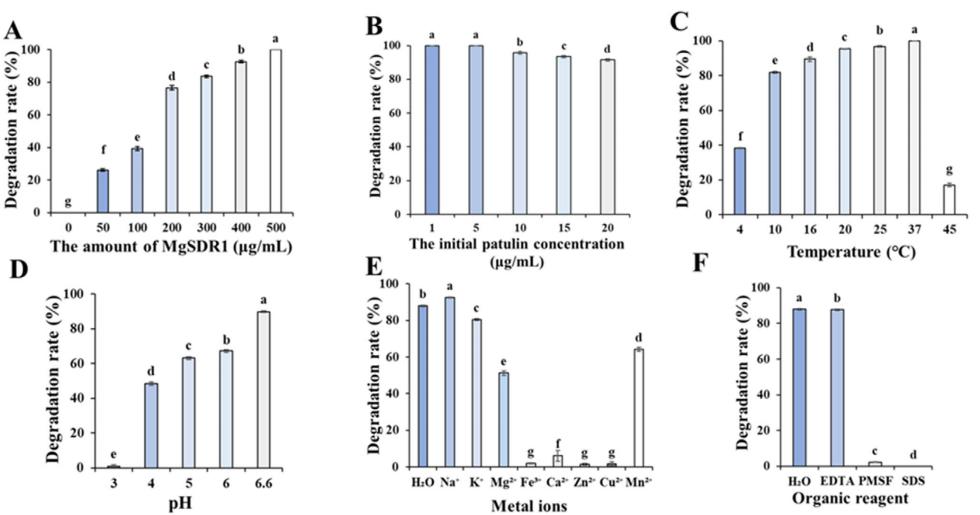
温度、pH、MgSDR1浓度和初始PAT浓度对生物降解过程影响较大。 图6A显示,MgSDR1浓度与PAT的降解率呈正相关。 当MgSDR1浓度为500 μgmL−1时,PAT的降解率为100%。 图6B显示,当初始浓度为1 - 5 μgmL−1时,PAT在相同的时间内完全降解。 而当PAT初始浓度为20 μgmL−1时,2 h内的降解率为91.44%。图6C显示,在25 ~ 37℃间,MgSDR1表现出最好的催化活性,完全降解率为100%。 图6D显示pH显著影响MgSDR1对PAT的降解速率。 其中,MgSDR1对PAT的降解率在pH 3.0时低于1%,在pH 4.0时达到48%,在pH 6.6时达到90%。
金属离子和有机化合物对酶的活性有显著影响。图6E显示,Fe3+、Cu2+、Ca2+、zn2 +的存在对MgSDR1的活性有较强的抑制作用,使其活性降低到10%以下。
k +、Mg2+和mn2 +存在时,酶活性保持在50%以上。 相反,Na+可以促进MgSDR1对PAT的降解。 图6F显示,PMSF和SDS显著抑制了mgsdr1介导的PAT降解,而EDTA对mgsdr1活性的影响很小。
鲜梨汁中PAT降解试验及品质测定
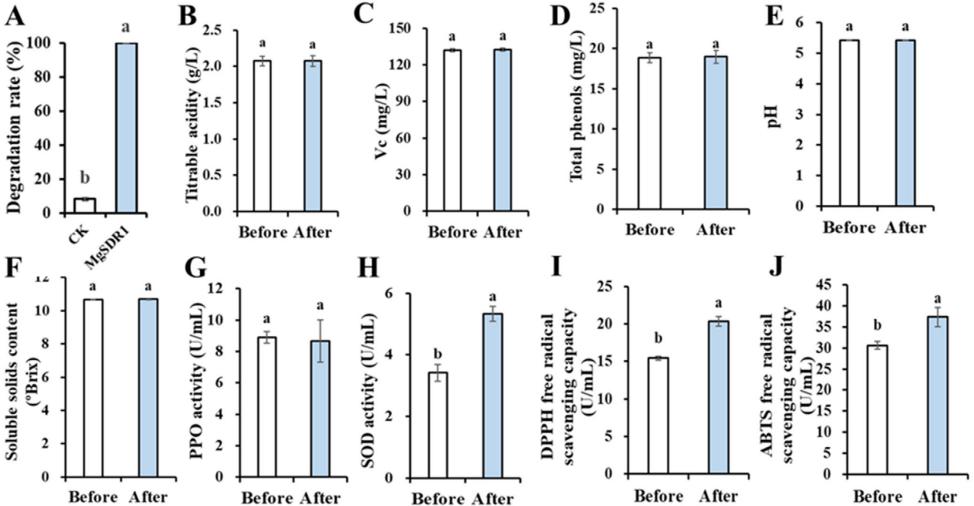
在鲜梨汁中验证了MgSDR1对PAT的降解作用。 新鲜梨汁中的PAT (10 μgmL−1)在12 h内被完全降解,降解率为100%(图7A)。 鉴于MgSDR1对鲜梨汁中PAT的降解效率较高,具有广阔的应用前景,作者对鲜梨汁的品质参数进行了测定。 生物降解过程对pH、多酚氧化酶(PPO)活性、可滴定酸度、维生素C (Vc)、可溶性固形物和总酚含量无显著影响(图7B-G)。 生物降解后,果汁超氧化物歧化酶(SOD)活性、2,2-二苯基-1-苦酰肼(DPPH)活性和2,2 '氮基-双-(3-乙基苯并噻唑啉-6-磺酸(ABTS)自由基清除能力显著提高。 上述结果表明,生物降解过程可以提高抗氧化能力(图7H-J)。 新鲜的梨汁呈黄褐色。 生物降解前后样品的颜色参数(L、a、b)、总色差(ΔE)、浊度、褐变指数保持一致。 这表明PAT降解过程没有改变新鲜梨汁的颜色、浑浊度或褐变程度。
采用气相色谱/质谱联用技术(GC/MS)测定生物降解前后香气成分的变化。 GC/MS结果显示,己醛、1-己醇、3-己烯-1-醇、乙酸、(Z)-、乙酸、己酯、癸醛含量均有所增加,壬醛和十二醇含量略有下降。 总的来说,生物降解过程促进了酯类、醛类和醇类的形成。
基因定点突变
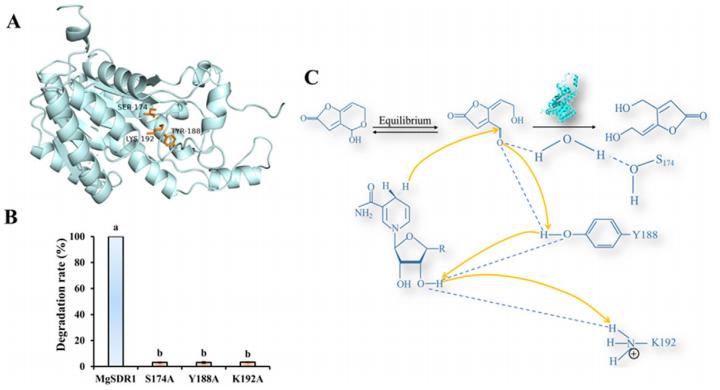
MgSDR1、MgSDR2和MgSDR3具有保守的催化三联体(图8A)。 通过位点诱变研究了关键催化三元组在MgSDR1降解PAT中的作用。 MgSDR1的活性位点(Ser174、Tyr188和Lys192)被Ala取代。 结果,突变蛋白(MgSDR1S174A、MgSDR1Y188A和MgSDR1K192A)的PAT降解率显著降低,活性几乎完全丧失(图8B)。 基于这些发现,作者提出了PAT降解的机制。 Tyr侧链作为催化碱,催化碱羟基的pKa可与NADPH的核糖羟基、带正电的赖氨酸和催化丝氨酸相互作用而降低。 此外,催化Ser的侧链可能与底物PAT相互作用,从而将其稳定在碱策略的结合口袋内的合理构象中。 催化产物反应开始于质子从Tyr的羟基转移到PAT的羰基氧原子。 随后,NADPH的氢原子被转移到羰基碳上,导致PAT的羰基转化为羟基(图8C)。